| Revised on November 19th, 1999 | ||||||||||||||||||||||||||||||||
|
器官形成研究グループ のホームページだ! |
||||||||||||||||||||||||||||||||
|
器官形成時の組織間相互作用 における 細胞接着因子と細胞間マトリックス |
||||||||||||||||||||||||||||||||
| はじめに | ||||||||||||||||||||||||||||||||
|
今から50年も前の1950年代にGrobsteinが、胎仔期の器官形成過程の「上皮ー間充織相互作用」において細胞間マトリックス成分がカギを握っていると指摘していたが、近年になってマトリックスの重要性が認識され、上皮の形態形成期におけるマトリックス成分の研究は徐々に活発になりつつある。われわれの最終目標は、器官形成時の組織間相互作用(Tissue
Interactions)とは如何なるものか、また組織間相互作用を媒介すると考えられるマトリックス成分とその働きを明らかにすることである。言い換えれば、組織間相互作用という概念を具体化することである。 一方、上皮組織の形態変化には細胞接着因子も重要な位置を占めていることも予想され、その変化も「上皮ー間充織相互作用」の影響下にあることも近年明らかになってきた。 そのために我々は、比較的よく研究され、取扱いが易しいマウス胎仔の唾液腺、肺臓などを対象として選んできたが、近年これらに加えて毛、乳腺、そして歯なども研究対象としている。 |
||||||||||||||||||||||||||||||||
| マウス胎仔唾液腺の発生 | ||||||||||||||||||||||||||||||||
|
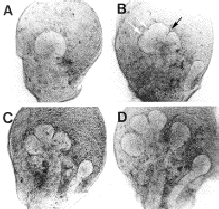
マウス胎仔下顎唾液腺(顎下腺、Submandibular
Gland)の形成は、妊娠11日目に口腔上皮が間充織中に陥入することに始まる。12日目になると上皮細胞は増殖を繰り返して電球状になり(下図-A)、その直後から上皮組織の分枝(epithelial
branching)が開始される。すなわち、12日目後期になると上皮の1か所から数か所に窪みが出来始め(下図-B)、その内のいくつかが13日目の初期には深く狭いクレフトとなる(下図-C)。そして13日目中期から後期にはこのクレフトは広く深い形になり(下図-D)、こうして出来た新しい小葉(lobule、上皮細胞のかたまり)は以後さらに増殖と分枝を繰り返してブドウの房状の唾液腺となる。一方、上皮を取り囲む間充織は、12日目の初期までは他の間充織との間に明瞭な境界線を持たないが、12日目後期にはその区別がはっきりした唾液腺間充織の形態形成が行われる。このことは上皮の形態形成と関連していると思われるが、これについては全く不明である。 |
||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||
| マウス胎仔唾液腺の発生。それぞれ12日目中期(A)、12日目後期(B)、13日目初期(C)、13日目中期(D)の連続写真を示す。Bでは明らかにクレフトになるもの(黒い矢印)と、どちらかがクレフトになると思われる2つの窪み(白い矢印)がある。どちらかが結果的には選択されていることが分かる。 | ||||||||||||||||||||||||||||||||
| 上皮形態形成の細胞間マトリックス成分による制御 | ||||||||||||||||||||||||||||||||
|
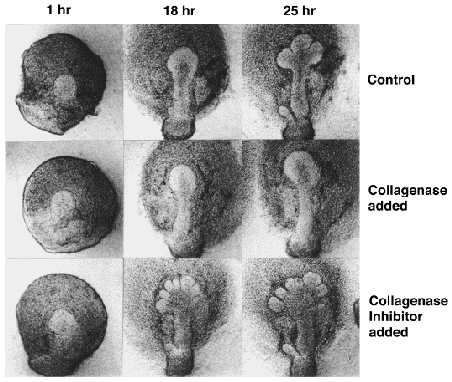
これまでの研究から、特に初期クレフトの形成にはマトリックス成分であるコラーゲンや現在広く知られているヘパラン硫酸プロテオグリカンなどが決定的な役割を果たしていることが明らかになってきた。なお、このプロテオグリカンは上皮細胞の増殖にも関係していることが予想されている。 例えば下の写真のように、コラーゲンの分解を抑えるコラゲナーゼインヒビター(現在はTIMP-1と呼ばれる)を培養液に加えると(下段)、クレフト形成が著しく促進されるのに対し、コラーゲンの分解するコラゲナーゼを加えると(中段)クレフト形成が全く抑えられる。 さらに、基底膜や細胞膜に存在すると思われるヘパラン硫酸プロテオグリカンのヘパラン硫酸鎖を分解する酵素であるヘパリチナーゼ Iを加えると上皮の正常な形態変化が阻害され、分解活性の弱いヘパリチナーゼ IIではその影響が弱いことが証明されている。 |
||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||
| ( Scott F. Gilbert著、"Developmental Biology" 5th Edition, pp. 684-686, 1997 および Y. Nakanishi et al., Developmental Biology, 113, 201-206, 1986 を参照) | ||||||||||||||||||||||||||||||||
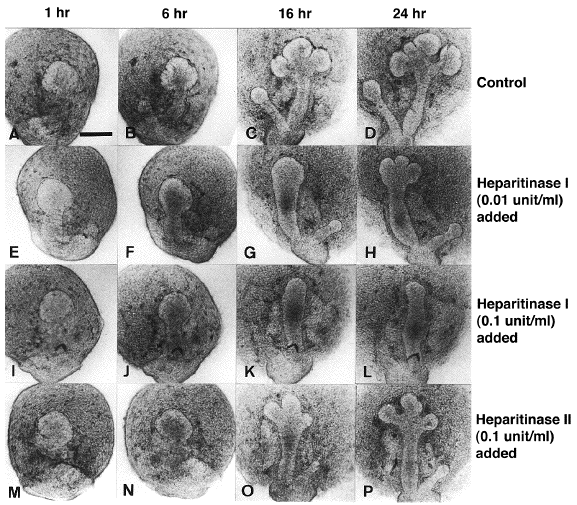 (Nakanishi et al., Develop. Growth & Differ., 35, 371-384, 1993 を参照) |
||||||||||||||||||||||||||||||||
| また、間充織が上皮に対してEGF-likeの成長因子を供給していることも知られているが、その実態は明らかでなく、上皮細胞と間充織細胞との直接接触による情報交換も考えられる。一方、初期形態形成時には活発な間充織の運動が観察されている(形態形成運動のアニメーションにリンク)。 | ||||||||||||||||||||||||||||||||
| 以上のような研究結果から、特にコラーゲンの関与を中心として新しい仮説を提唱した。 | ||||||||||||||||||||||||||||||||

|
||||||||||||||||||||||||||||||||
|
その後の展開−器官形成初期過程における 上皮の組織構築の激変 |
||||||||||||||||||||||||||||||||
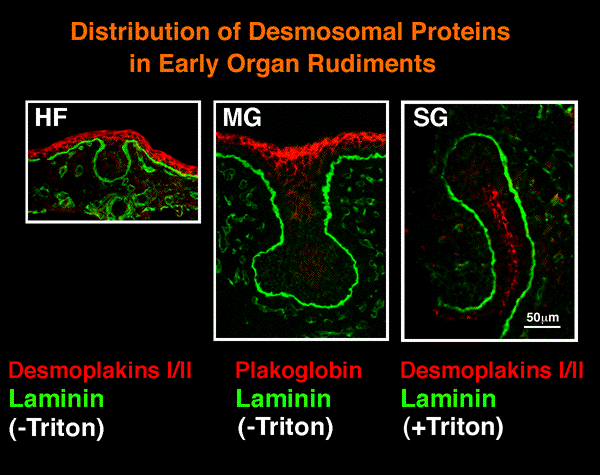
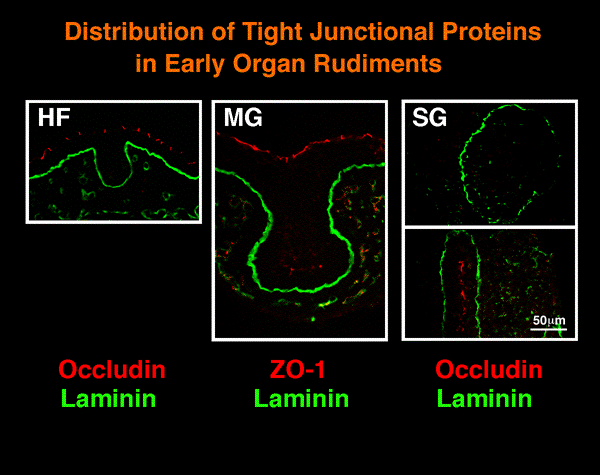
| 驚いたことに、桧枝らは最近上皮細胞の活発な運動をビデオ画像として観察したことから、器官形成初期の上皮組織の性格をめぐって新たな展開が始まっている。例えば、デスモソームやタイトジャンクションの構成分子であるデスモプラキンやオクルーディンは初期の上皮塊には存在せず、それは典型的な上皮とは著しく異なることが証明された。同様のことは、唾液腺のみならず毛包や乳腺の形成初期にも起こっていることが明らかになりつつあり、その生理的意義−上皮の形態形成と細胞分化−に関して新しい視点が必要になりつつある。 | ||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||
| 唾液腺(SG)、毛包(HF)および乳腺(MG)の初期budにはデスモソームやタイトジャンクション関連の接着関連分子が検出できないことに注意 | ||||||||||||||||||||||||||||||||
|
このように、唾液腺の初期クレフト形成に絞って始めた解析であるが、現在はその対象が毛包や乳腺にも広がり、対象となる分子や生理現象もさらに広がる情勢となっている。 |
||||||||||||||||||||||||||||||||

|
||||||||||||||||||||||||||||||||
| なお、関連論文、related paper、はここからリンク。 | ||||||||||||||||||||||||||||||||

|
||||||||||||||||||||||||||||||||