
| |||||

|
|||||
|
|||||

|
|||||

|
|||||

|
|||||

|
|||||
|
|||||

|
|||||

|
|||||
|
|||||
|
|||||

|
|||||
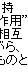
|
|||||

|
|||||

|
|||||
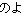
|
|||||
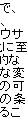
|
|||||

|
|||||
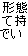
|
|||||
|
|||||
|
|||||
|
|||||

|
|||||

|
|||||
|
|||||

|
|||||
| III−4.”相互作用”の限界と”構造的行きづまり” | |||||
| それでは相互作用は際限なく続くものと考えてよいであろうか。しかし細胞の分裂から次の分裂、または環境による分裂停止、さらに大きな事としては細胞、組織あるいは個体の死が存在するからには、この”相互作用”の中に何らかのかたちで限界を持たなければならないことは明らかであり、それが規定されない限り多くの矛盾を含んでくる。 | |||||
| ここで・−2で述べた細胞外物質の概念規定を思い出していただきたい。そこで我々は”非常に大きな構造変化の可能性を含んでいなければならない”と規定している。つまり、構造は生合成の過程においても、後からの修飾においても容易に変化しうるが、しかしその変化は”有限”である。たとえば、最も考えやすいコンドロイチン硫酸蛋白複合体を考えてみよう。この分子の糖鎖は蛋白に結合しているが、その siteには限りがあり(セリンの数には限りがある)、また長さも蛋白との関係において規定される。また糖鎖中の硫酸基の数は、特殊な場合を除きその糖鎖の長さと官能基の数以上にはなり得ないことは明らかであろう。 | |||||
| この観点で図1を見直すとひとつ重要な点が存在する。それはコンドロイチンタイプといわれる硫酸基のないN-アセチルガラクトサミン残基の量が胎児期の中期から終わりにかけて徐々に減少して行くことである。その時の細胞がアセチルガラクトサミンの4位と6位に硫酸基を導入する2つの酵素しか持たないならば、この相互作用は胎児期の末期に構造的な変化の限界、つまり”構造的行きづまり”を実現してしまう。軟骨細胞は、さらにこの限界を乗り越える形であたらなケラト硫酸の合成を開始する。この分子もコンドロイチン硫酸蛋白複合体同様にその微細な変化を観察すれば、たとえば硫酸含量の少ないものから多いものへと変化していると考えても不思議ではない。しかし、40才位のところからほとんで変動がないことは、軟骨組織として完全に限界に到達し、それ以上の新たな情報を引き出せないことを示している。 | |||||
| これまでの議論の中で重要なことは次のことである。すなわち、我々が扱おうとしてきた”相互作用”は、その構造上の裏付けとして、コンドロイチン硫酸蛋白複合体の場合、糖鎖の数としては少ないものから多いもの、すなわち分子としては小さいものから大きいものへ(Hardinghamら)、また硫酸含量としては少ないものから多いものへ(図1)、そして糖鎖の長さとしては短いものから長いものへ(木全ら)と変化していることである。このことは”量”ということで一般化したとき、小から大への一方向の変化しかないことを示しており、少し飛躍するが、たとえば個体は小さいものから大きいものへとしか変化しないことと一致して興味深い。 | |||||
| III−5.”構造的行きづまり”とその生物学的意味 | |||||
| いままで扱ってきた軟骨のデータ(図1)でこの両者の関連をみてみたい。まず、胎児期の中期から終わりにかけてCタイプとコンドロイチンタイプの合成が減少し、これがAタイプの増加と一致する。全てのアセチルガラクトサミンの4位と6位がふさがれて硫酸の入れる位置が無くなるとという限界に近づくと、それを乗り越える形でケラト硫酸の合成が行われる。この変化が定常状態になったとき、その組織としての限界に到達していると考えられるが、しかしこの場合新しい細胞外物質を作り出す情報を遺伝子の中から引き出せないことを示している。すなわち、アセチルガラクトサミン残基が硫酸残基によって修飾されたものを作る過程が原基形成であり、新しくケラト硫酸合成による”相互作用”の克服過程がAdultの軟骨組織への”成長”過程である。しかしながらケラト硫酸の構造が変化の可能性を失って固定化したものになり、しかもこれに情報の有限性が関係したとき、その組織は次第に”老化”して他の細胞にとって代わられるか、組織が崩壊するかのどちらかとなってしまう。だか、なぜ”老化”するかはわからないが*、いまのところ合成抑制による細胞活性の低下としか言い得ない。 | |||||
| すなわち、”成長”とは、細胞と細胞外物質との相互作用の結果として構造的に行きづまるまでの過程であり、その中には組織の急変はないが、たとえばケラト硫酸の新たな合成という、ある意味では”分化”の概念をも含んでいる場合も存在する。この成長過程の把握で重要なことは、相互作用の結果とDNAの情報の有限性の中で必ず”構造的行きづまり”を実現することであり、このことが同時に老化を必然ならしむることである。すなわち、”成長”と”老化”は細胞と細胞外物質との”相互作用”の結果としての、化学構造の一方向への変化の表現に他ならない。これを一応個体に総体化したときには、個体の死は必然であり、だからこそ新たな個体の生成が”生殖”として必然化されるのである。 | |||||
| *ひとつは、分解しにくいものを作ってしまうところにあると言える。たとえばケラト硫酸を分解する酵素系は未だ動物では見つからない。これはTemplateの問題としても、コンドロイチン硫酸の場合、これを分解すると言われるヒアルノニダーゼは、硫酸基の無いものの方が硫酸基のあるものより15倍も効率よく分解することが知られているので、”相互作用”の結果の産物は徐々に分解しにくいものを作ってゆくことになると考えられる。 | |||||
| III−6.”分化”の概念と”構造的行きづまり” | |||||
| このようにみてくると一応成長と老化は、”構造的行きづまり”という視点の中で一貫して取り扱うことが可能と考えられるが、この概念の一般性を検証するためにもたとえば分化の問題をこの範疇で取り扱えないかを検討してみたい。 | |||||
| 現在の時点では分化した細胞の性質として特異性の相互排除、たとえばアルブミン合成の特異性とグロブリン合成の特異性は相容れないことが明らかになっていると言われる(57)。つまり分化の過程は、特異性の獲得ということだけではなく、むしろ一般性としては、他の特異性の排除を含むものでなければならないとされている。すなわち、発生から分化の過程は、一個の受精卵という全ての情報の発現可能なものから情報発現の可能な量が減少してゆくことであると言い換えることができる。このことはひとまとめにして次のように言えないだろうか。まず相互に排除しあう遺伝子は何らかの構造的な関連性があり、情報の発現という”相互作用”を通して排除される遺伝子の環境に構造的な変化が生じ、それの繰り返しの結果として構造的な限界に到達してしまう。そしてこの結果としてその遺伝子は排除されてしまう。もしそうだとすると、”構造的行きづまり”の視点から”分化”を規定することが可能となってくる。 | |||||
| ここで現在情報発現にとって重要だといわれるヒストンを考えてみよう。現在まで数種類のヒストンの一次構造が決定されており、ある種のヒストンの構造をウシとエンドウで比較するとアミノ酸配列の違いはわずか2カ所で、それはDNAの塩基一個の変化で説明できるほど厳密なものであるといわれている(58)。しかしながらヒストンには分画によって数多くのフラクションに分かれ、種特異性や組織特異性のあることが明らかにされている。もちろん、一次構造の違いによる多種類のヒストンの存在は当然であるが、ここで問題にしたいのは、それの修飾反応である。たとえば、ある種の高Lys, Ser型で、子ウシ胸腺からとったヒストンは、125個のアミノ酸のうち、側鎖修飾可能なアミノ酸はLys 20個、His 3個、Arg 8個、Ser 14個、Thr 8個で合計53個となり、約40%のアミノ酸は修飾を受ける可能性を持っている(59)。もちろん、ヒストンの塩基性はこのうちのLys, Argの構造が表現するものであり、他のヒストンの場合についても本質的には同じであろう。 | |||||
| いま、DNAからの情報発現における相互作用を、細胞と細胞外物質とのそれと同じように何らかの構造変化をその基盤に持っているとすれば、細胞外物質の場合と同じように、相互作用をする系の中の物質に対して構造変化の可能性という観点を持ち込まざるを得ない。このような観点から見ると、”核内相互作用物質”として最も考えやすいのは先程述べたヒストンであろう(もちろんこれ以外のものもあり得る、修飾可能なRNAとか)。あるヒストンは約40%の修飾可能なアミノ酸を持っていることを述べたが、たとえば次のような修飾反応がある(60)。Lysineはε-N-acetyllysine、ε-N-monomethyllysine、ε-N-dime thyllysine、そしてε-N-trimethyllysineに、Histidineは3-methyl-histidineに、Arginineはω-N-monom ethylarginineとα-N-methylguanidinomethylated arginineに、そしてSerineはphosphoserineにそれぞれ変化を受けることが知られており、実際にはもっと多くの修飾の可能性が示唆されている(60)。これらのことは軟骨細胞の細胞外物質である酸性ムコ多糖の構造変化の様子と極似しており、”相互排除”という”分化”をその遺伝子の環境の不可逆的な”構造的行きづまり”として理解できるようである(いまはその例としてヒストンをあげた)。 | |||||
| 我々はここまで、細胞と細胞外物質との相互作用もまた核内におけるDNA、ヒストンそしてこれに絡むであろう他の因子との相互作用も同じものとして扱ってきた。しかし前者は、ある特異性を持った情報の発現は変化させず、合成活性発現のレベルでその分子内における量的な変化を通じて質的な変化をもたらすという意味で”成長”、”老化”の問題として考えてきた。これは”弱い相互作用”と呼ぶべきかもしれない。それでは後者の分化の場合はどのような相互作用として規定したらよいのだろうか。特にこの場合重要なことは、分化はその生物学的時間の中で厳密に行われているということである。そうすると分化とは、たとえば二重のチョックシステム、言い換えればその細胞と細胞外物質との相互作用の限界性と、その時までの情報発現の限界性という、いわば二重のものを克服するという”強い相互作用”として考えてはどうであろうか。つまり、2つのphaseが一致したときに新しい情報を、たとえば他の細胞が作っている細胞外物質を媒介にして新しい蛋白(酵素)として引き出し、相互の限界を克服するのである。 | |||||
| 一方、分裂という過程においてもヒストンなどのものをDNAから切り離し、DNAの複製を可能にすることは予想されるが、この際ヒストンが何らかの修飾を受けるのであろう。そしてこの繰り返しがヒストンに”構造的行きづまり”をもたらし、加齢とともに分裂能力の低下をもたらしてしまう。 | |||||
| このような相互作用をいろいろな意味に解釈することは可能であるが、その基本は種々の側鎖の形成が行われることにあり、またDNAという酸性物質と相互作用しなければならないという、いわば二重の意味で塩基性であることがヒストンには要求される(もちろん多くの人がこの点に言及している)。そしてこの相互作用がきわめて多様に、しかも連続的に行えるように細胞外物質と同じような構造上の可変性を備えていると見るべきだろう。ただヒストンが細胞外物質と異なるところは、後者は蛋白以外の部分は可変である、たとえば長ささえも変えられるのに対して、前者の場合はその長さは厳密に決定されている点である。ここでは主にヒストンのことについて述べたが、この系の中にあるDNAもこの範疇の対象であるかもしれない。なぜなら、DNAのメチル化酵素は厳然として存在するからである(61)。 | |||||
| なお、分化を相互排除という観点をとり、それによって図1をみた場合、ケラト硫酸の増加に伴ってコンドロイチン硫酸Aがそれに対応して減少してくることは、この2つの特異性が相容れないものとして存在しているものであると理解される。 | |||||
| III−7.”構造の自由度”と細胞構造 | |||||
| このようにみてくると、細胞というものをひとつの”構造の自由度”という観点からもみることが可能となる。すなわち、細胞の中心には構造の自由度の小さいDNAが存在し、それを取り巻くヒストンはTemplate-d ependentではあるが側鎖の修飾による構造変化の可能性を持ち、細胞膜あるいは細胞外にはその鎖の長ささえ可変であり、なおかつ側鎖の修飾さえも可能である、ある意味では最大の構造の自由度を持ったムコ多糖蛋白複合体あるいは糖蛋白質などが存在する。そしてその間には、細胞質として行動の自由度を持った分子または”積木の化学”の可能なオルガネラ(10)が介在する。 | |||||
| これをみると、細胞というのは中心から外へ三次元的な空間的拡がりを持つと同時に、明らかに構造変化の自由度が内から外へ拡散する方向があるものである。逆に言えば、細胞は内から外に向かってより構造に自由度を持つ物質によって構成されていることによって空間的な拡がりが保証されているのである。我々はふだん、細胞は外から情報を得ることが重要であるといったりするが、この必然的な内容は、実は細胞のでき方それ自体の問題であり、細胞外や膜の化学構造あるいは三次元構造の変化を媒介にしてはじめて逆の方向、すなわち外から内への情報伝達が可能になるのである。このことについてはこれ以上述べられないが、細胞と環境との相互作用や情報の獲得形式の問題である進化の問題を扱う際には重要視しなければならないであろう。 | |||||
| III−8.”構造的行きづまり”と細胞分裂 | |||||
| 今まで述べてきたことは、活発な相互作用の過程は”成長”であり、”構造的行きづまり”になってしまって固定化した状態が”老化”である。そしてこの行きづまりを打開できたのが”分化”と規定した。各組織でそれぞれに分化の極に到達してしまった次に来るのが個体の死であり、それ故に個体発生が必然化されるのであろうということであった。 | |||||
| この問題の次元を少し下げて細胞分裂をこの延長線上で把握するとどうなるであろうか。そうすると当然のことならが細胞分裂は、細胞あるいはそれを取り巻く環境の中にいる細胞の何らかの限界を突破するものと考えなければならない。たとえば限界に来た細胞外物質を、分裂に際しての新たな合成で稀釈することに分裂の意味があると思える。もちろん、細胞の容積の限界を克服する方向もあるであろう。このことは核内における相互作用の限界の打開としても同じように言えるであろう。つまり、ヒストンの構造的な行きづまりをも同じように打開する。しかし往々にして、動物細胞の分裂回数には明らかに限界があることが指摘されている。この点については・-6で述べたように、分裂を行うためのDNA複製に対して行われるヒストンの修飾結果が、その回数を重ねる度に蓄積し続けて完全な構造的行きづまり状態になることとして理解される。もちろん、どちらの場合にも分解酵素系による限界の打開は、組織や細胞の代謝活性の維持あるいは増進に有効であるが、次のような問題を含むものである。 | |||||
| ここであえて我々が分解酵素なるものを持ち出したのは、いずれ分解酵素についての規定が必要だからである。もし、相互作用の結果として生じる変化した構造を分解酵素系が完全に元に戻してしまう、あるいは、分裂などの際に構造的な限界を分解酵素系がやはり完全に元に戻してしまえるものとしてそれを規定したとするならば、相互作用を変化の源泉として捉えようとすることははじめから存在し得ないことである。つまり、我々が最初相互作用の概念を規定したとき(III-1)、すでにその中に相互作用の結果生じた変化した構造を完全に元に戻してしまうよな分解酵素系を、細胞あるいは組織が持つことは認められないという立場が存在したのである。 | |||||
| 逆に、このような系が存在するひとつの例がバクテリアともいえる。バクテリアの分裂は、たとえば前の分裂から次の分裂までの連続的な細胞壁(膜)成分の変化の結果生じた構造的な限界を分裂によって克服する、そうゆう意味を持つと考えると、ここでは唯の稀釈ではなく、限界にある状態の完全な解除が行われたと考えなければ無限といえるほどの分裂は行えないはずである。むしろこのようなことが行いうるところに、バクテリアは一個が全体として存在しうる必然性があるのかもしれない。これと同じような系が癌細胞であるともいえるであろう。 | |||||
| ところで、多細胞生物系で分解酵素なるものを上のように規定したからといって軽視するつもりは全くなく、逆により重要なものとして把握される。話をより高次な血清のレベルに戻してみよう。血清は個体の全てに行き渡る点において各組織をバランスを保って維持するシステムと考えてよいであろう。いままでの議論の延長線上当然のことであり、幾つかの事実としても明らかにされている(62, 63, 64)。また各組織が完全に分化しきったとき以降に持つ血清の役割は特に老化との関連で重要であろう。なぜなら血清の持つ分解酵素(65, 66)に注目したいからである。少なくともこれまでの議論において、”構造的行きづまり”を細胞内外(分裂の限界を含む)に持ってしまった組織を維持するには、相互作用が可能になるように、常にある程度の、合成を越えない、限界状態の解除が必要だからである。もちろん、細胞内外を問わずである。このような視点から、血清中の分解酵素系をただ組織から漏れ出たものとしてではなく、積極的に組織ひいては個体維持に必要なものとして把握し直すことが大切である。しかし、所詮血清成分も限界に到達するであろう細胞の生産物以上にはなり得ない。 | |||||
| いままでは、”構造的行きづまり”を分裂や分解でいかに突破するかの問題であったが、その前に特殊な化学構造でその限界を乗り越えている組織の変化を起こさない方向があるはずである。その例が、いわゆる永久軟骨であるイカの軟骨のコンドロイチン硫酸Eタイプ(67)ややはり永久軟骨であるサメにみられるケラトポリ硫酸(68,69)である。Eタイプというのはコンドロイチン硫酸AのN-アセチルガラクトサミン-4-硫酸残基の6位にもう1モルの硫酸基を持つものであり、しかも糖鎖の長さとしては新しいグルコース残基を糖鎖の中に導入することでより高分子を実現しているらしい(70)。またサメのケラトポリ硫酸は、普通のケラト硫酸の硫酸基は繰り返し2糖単位あたり1モルなのに対して1.8モルと高い。これもひとつの克服の方法であろう、そしてそれによって組織の限界を遠ざけ、たとえば永久軟骨として実現していると考えられる。また、細胞と細胞の関係によって限界を克服している例はたとえば神経細胞であろう。分裂しない神経細胞が分裂するグリア細胞によって取り囲まれているのは(71)、神経細胞の内外に存在する限界状態が回りに存在するグリア細胞によって解除されるためであろうと思われる。以上、2、3の例を挙げたが、相互作用物質である酸性ムコ多糖の構造は枚挙にいとまがないほど多種多様存在していることが予想される。 | |||||
| III−9.”分化”した細胞核から”全能”なる細胞核への逆戻り | |||||
| ここにひとつの問題がある。オタマジャクシの腸の細胞として完全に分化しきった核を取り出し、これを卵細胞の核と入れ換えると、その核の機能は卵細胞の核と同じ全能の核に戻る現象がある(72)。いままでの内容からこの現象を考えてみると、核内の”構造的行きづまり”(いまの場合ヒストンでよい)が細胞質との関係で、たとえば分解酵素系によって解除されたと理解される。これが卵の細胞質の特殊性であって、ある程度の卵割まではその細胞の核が限界を来さないような、たとえば分解酵素系の質と量があらかじめ用意されていると考えられる。この点の問題が、実は卵細胞が成熟してくる不等分裂の過程に含まれるであろう。言い換えれば、バクテリアのように一個の細胞になる過程である。この「戻り現象」にはかなりの時間が必要であり、移植された核の障害の何らかの解除が進められた可能性が高い。 | |||||
| III−10.細胞と細胞外物質の相互作用はどのように行われるか | |||||
| いままで述べてきたことは一貫して、相互作用が変化を生む元であるということであった。それではそのようなことがどうして可能であるのか。現在全くblack box出あるが、それを指し示すようなデータがないわけではない。そこで、いままでのことを思考するきっかけとなったデータを少し再編してみよう。 | |||||
| たとえばAshwell一派の実験である(30, 31)。一般に血清糖蛋白質はシアル酸を末端として持つのであるが、たとえばceruloplasminを静脈に注射すると時間のオーダーで血液中を循環しているが、たとえば10個持っているシアル酸残基のうち2個ほどのシアル酸を除いたものを静脈に注射すると分のオーダーで肝臓の膜を介して取り込まれ分解されてしまう。このことは2個のシアル酸を除くことで膜に対して物性的に同質化し得たと解釈できる。なぜなら膜の糖蛋白質はほとんど血清糖蛋白質とよく似た組成であるからである。さらにこのような同質化を合成分泌過程の裏返しとみれば、その過程には膜に存在するシアル酸を持つ糖蛋白質に対し、出てゆくce ruloplasminがある程度以上のシアル酸を持たないと細胞外へ放出されないと考えられ、分泌には膜に対する異質化、つまり膜にあるものに対して異なったものへ転化することが必要と考えられる。 | |||||
| 一方、II-2の図4、5で述べたKern, Swensonら(21)のリンパ細胞によるγ-グロブリンの合成分泌過程は、詳しくは述べないが簡単にいえば次のようにいえる。糖部分の最後の糖であるシアル酸が付くまではそのγ-グロブリンは細胞内に存在するが、いったんシアル酸が結合すると速やかに細胞外に分泌される。ここから少し飛躍するが、糖部分の蛋白部分への添加は膜への物性としての同質化であり、Ashwellらのデータとかみ合わせると、結合しうる部分にほぼ全てシアル酸が結合することは異質化の過程だと考えられる。このような事実から我々が概念として導き出したことは、膜を通しての分泌には、分泌されるものの合成過程の中に膜に対しての何らかの意味で同質化と異質化の過程があって、この異質化のところに細胞外物質の多様性を実現するカギがあると思われる。なぜなら、培養細胞の細胞膜にコンドロイチン硫酸が存在するといわれ(52, 53)、また膜の物質を膜と接触しうる細胞外物質とおくこともできると考えれば、それと異質化することを化学構造の中で考えれば常に異なるものを合成しなければ分泌が行えず、合成活性も抑えられるであろう。ここの”老化”の契機がある。・-3で引用したNevo, Dorfmanの実験はこのことまでも表現しているのではないだろうか。この場合、細胞外に加えたコンドロイチン硫酸は相互作用のきっかけを与え、合成分泌を引き起こす水先案内人のような役割を果たしているとみられる。このようなことが分化の契機であることをStrudelが指摘している(73, 74)。 | |||||
| この項の中で重要なことは、細胞と細胞外物質との”相互作用”は、細胞外物質が合成分泌される過程の問題であり、それは多分分泌が決定される膜との”関係”にあるということである。そしてこの”関係”とは、単純な酵素と基質の問題とは考えにくく、むしろすでに合成していた同種の化合物やその他の因子を含めた”鋳型”としての膜構造の問題であろう。 | |||||
| このようなこと考える結果として、分泌されて細胞外に出たものが逆にその細胞を規定するような分子の分泌過程では、その分子と膜との相互作用を必然的な内容として持ち、なんらの細胞への規定性を持たないものの合成分泌は、その分子と膜との相互作用は本質的な意味において存在しないと考えられる。たとえば、膵臓のアミラーゼのように消化器官内への分泌物質は、その合成過程で完全に顆粒状に包まれ、分泌される時にはその顆粒の膜と細胞膜との接触によって中のアミラーゼは放出されてしまう(75)。このようなかたちで様々な物質の分泌のされ方を規定し、分類し直すことも可能であろう。 | |||||
| III−11.まとめ | |||||
| 我々は細胞外物質の構造の多様性を観る視点を作り出すために、相互作用という言葉の意味を把握し直し、それを用いて”構造的行きづまり”という概念を引き出してきた。そしてこの概念を用いて”分化”、”成長”、”老化”という一連の生体変化を見直そうとし、それが可能のように見える。つまり、発生したものは必ず老化するという多細胞生物の持つ一方向の変化−生物的時間−は相互作用の過程の中で、鋳型によらないで合成される分子構造の中に蓄積されてゆく過程の産物であると考えられる。しかし、あくまで我々は、その変化のきっかけ(契機)を述べただけであって、たとえば相互作用の限界がなぜ老化という形態をとるのかは全く想像の域を出ない。 | |||||
| またIII-1で述べた酵素の問題を考えると、酵素反応という相互作用の中で酵素は変化しないというのがいわゆる酵素学であるが、本当にそうであろうかという素朴な疑問がでてくる。つまり酵素が現在金属イオンのようなものでなく、まさに酵素であるとは一体どうゆうことなのかという問題であり、Sabato, Kaplanら(76)は、この点を意識していたように見えるが現在全く分からないとしかいいようがない。 | |||||
| いずれにしても、この・章に書いた内容の限界がよく理解できない。それは我々が日頃慣れ親しんでいる実体主義的な取り扱いでない概念的なものであることに由来しているようである。そしてまた、I章で分子生物学的決定論なるものを概念として徹底的に批判しえてはおらず、ただ結果的には分子生物学の弱点を突いたに過ぎないことが一連の論理の限界の不明確さをもたらしているのだろうと思われる。我々は、確かに分子生物学の克服を目標としたが、未だ対置に過ぎないことは十分認識しておく必要がある。 | |||||
| あとがき | |||||
| 書き終えてよくよんでみると、読みにくいことはもちろん、実にいろんなことに口を出したということを感じる。これを読まれた方は、我々を実に恥知らずな奴等と思われるかもしれない。しかし、生物の持つ時間とか場所の問題が我々の手から抜け落ちていることも事実であり、それを扱う議論の材料のつもりであって、アイディア競争をするつもりのないことは申し添えておきたいと思う。 | |||||
| 1972年12月 中西康夫、Y.K. | |||||
|
I.分子生物学の再検討とその立場の持つ困難性 (上のタイトルをクリックするとリンク) |
|||||
|
II.糖蛋白質の構造のHeterogeneityと糖ヌクレオチドからの糖転移反応 (上のタイトルをクリックするとリンク) |
|||||